
 Potential for Direct Nitrate-Nitrite Inhibition of Submerged Aquatic Vegetation (SAV) in Florida Springs: A Review and Synthesis of Current Literature
Potential for Direct Nitrate-Nitrite Inhibition of Submerged Aquatic Vegetation (SAV) in Florida Springs: A Review and Synthesis of Current Literature
Todd Z. Osborne1,2*, Robert A. Mattson3, Michael F. Coveney3
1 Whitney Laboratory for Marine Biosciences, University of Florida, St. Augustine, FL
2 Wetland Biogeochemistry Laboratory, Soil and Water Science Department, University of Florida, Gainesville, FL
3 Bureau of Environmental Sciences, St. Johns River Water Management District, Palatka, FL
*corresponding author: 9505 Ocean Shore Blvd., St. Augustine, Fl 32080 (904) 461-4000 osbornet@ufl.edu
Received May 7, 2015; Revised March 11, 2015; Accepted April 1, 2016; Published: March 7, 2017; Available online: March 8, 2017
Abstract
Current observations of water quality in groundwater discharge from springs in Florida show anthropogenic enrichment of nitrate plus nitrite (NOx-N) generally attributed to fertilizer application and/or wastewater or manure sources in individual spring sheds. Excessive levels of NOx-N have been implicated in eutrophication of, and observed changes in, submerged aquatic vegetation (SAV) communities in several spring runs. While the indirect effects of nitrogen (N) enrichment on SAV, are well-documented (i.e., algal productivity resulting in shading of macrophytes), there is considerably less information available concerning direct effects of NOx-N such as toxicity or inhibition of macrophyte growth. This manuscript constitutes a review of the pertinent literature and synthesis of the current understanding of elevated NOx-N in aquatic systems and the effects on SAV as viewed from the prevailing eutrophication paradigm, as well as, explores the hypothesis that NOx-N may have direct inhibitory effects on SAV growth in Florida springs.
Introduction
Phosphorus (P) is often considered the limiting nutrient in freshwater ecosystems, and thus when available in excess, it is implicated as a causative agent in eutrophication (common eutrophication paradigm). Nitrogen (N), however, has been traditionally viewed by many as a lesser contributor to eutrophication of freshwaters, either because of the over-shadowing nature of P issues or due to the ability of many cyanobacteria to fix atmospheric N, a process that significantly reduces perceived N limitation. This pre-vailing view stems from research conducted to elucidate the role of P in eutrophication (Schindler and Fee 1974; Schindler 1978) following the 1960s chemical industry claims of no effect of increased P in aquatic systems (Barker et al. 2008). More recently, several researchers have reasserted the view that N either alone or in concert with P, may exert ultimate control over algal productivity and subsequently macrophyte productivity in aquatic systems across the globe (Turpin 1991; Talling and Lemoalle 1998; Maberly et al. 2002; Clark and Baldwin 2002; James et al. 2003; James et al. 2005; Sagario et al. 2005; Dzialowski et al. 2005; Weyhenmeyer et al. 2007; Li et al. 2008). The extreme of this viewpoint suggests that N may have been the limiting nutrient in most northern hemisphere lakes and rivers prior to substantial N fertilizer utilization, which alleviated N limitation and catalyzed P limitation in enriched systems (Bergstrom and Jansson 2006). A recent meta-analysis of published nutrient limitation studies found the number of N limitation cases to equal those of P limitation (Elser et al. 2007) and a significant number of cases of co-limitation. Moss (1990) contends that co-limitation was the normal condition prior to anthropogenic enrichment of nutrients. Recently, the role of N enrichment in alteration of ecosystem health has received renewed attention in aquatic ecosystems (Porter et al. 2013; Baron et al. 2013).
Numerous studies of eutrophication of freshwater ecosystems have observed a catastrophic shift from macrophyte to phytoplankton dominance after anthropogenic increases in available P (Wetzel 2001; Lacoul and Freedman 2005; Reddy and DeLaune 2008). The process involves rapid utilization of excess nutrients by phytoplankton and epiphytic algae, which results in explosive algal growth. The shift in primary productivity is also self- reinforcing, as turbidity increases with algal productivity, light becomes limiting to submerged aquatic vegetation (SAV) (Burkholder et al. 1992; Van den Berg et al. 1999). Death and decomposition of SAV only exacerbates excessive nutrient conditions by mineralization of organic nutrients bound in plant tissues.
Often, N and P have a positive synergistic effect on phytoplankton productivity. In a study by Sagarario et al. (2005), N and P additions alone did not show a significant effect, but when combined, the increase in phytoplankton and epiphytic algal biomass was dramatic. In addition to light attenuation by phytoplankton proliferation, epiphyte biomass burden and subsequent shading can be a primary causal mechanism for SAV mortality under eutrophic conditions (Borum 1985). In many spring runs in Florida, proliferation of epiphytic algae, as well as benthic macroalgae, have been observed concomitantly with declines of SAV communities (Stevenson et al. 2004; Frazer et al. 2006; Pinowska et al. 2007; Stevenson et al. 2007; Quinlan et al. 2008; Brown et al. 2008). Water quality in groundwater discharged from many springs in Florida has shown significant increases in NOx-N concentration, attributed predominantly to fertilizer application and/or wastewater or manure sources in individual spring sheds (Jones et al. 1996; Katz 2004; Albertin et al. 2012). Odum (1957) reported mean NOx-N concentration of 0.45 mg L-1 for Silver Springs in the 1950s which had risen to over 1 mg L-1 by 2005 (Munch et al. 2006; Quinlan et al. 2008). More dramatically, Rainbow River’s NOx-N concentrations have increased from 0.08 to 1.22 mg L-1 (a 15 fold increase) over the last 50 years (Cowell and Dawes 2008). Interestingly, during this period of increasing NOx-N, P concentrations have remained constant (Maddox et al. 1992; Scott et al. 2004) suggesting N to be the limiting nutrient in these aquatic systems. Perhaps more interesting is that not all springs with increasing NOx-N concentrations are experiencing increased algal productivity (Heffernan et. Al 2010), but many are experiencing declines in SAV health. These observations, while somewhat inconsistent with the common eutrophication paradigm, have prompted several hypotheses as to the role of increased nitrogen availability in the observed loss of SAV in spring systems statewide.
This manuscript reviews these competing hypotheses concerning the relationship between elevated NOx-N and observed ecological changes, specifically declines in SAV coverage and increases in epiphytic and benthic algae, in regional spring ecosystems. Significant attention is given to potential inhibitory effects of elevated NOx-N on SAV growth in springs and possible mechanisms for this inhibition are discussed. These mechanisms focus on the assimilative nitrate reduction process, resulting buildup of toxic ammonia (NH3), and energetic consequences of unregulated NOx-N uptake on SAV. This discussion concludes with a call for research to clarify the role of elevated NOx-N in the observed degradation of SAV communities.
Competing Hypotheses
The initial hypothesis (H1) posited by members of the scientific community, as well as the general public, was that the increase in N availability, observed as NOx-N concentration in spring waters, alleviated N limitation and therefore was responsible for a shift in primary productivity from SAV to epiphytic algae and benthic macroalgae. While there have been studies to report N utilization by algal mats in Florida springs (Cowell and Botts 1994; Cowell and Dawes 2004; Albertin 2009; Sickman et al. 2009), there have been several observations that contradict the common eutrophication paradigm, namely the lack of significant increase in other forms of N (Cohen et al. 2007) or P (Maddox et al. 1992; Scott et al. 2004) in spring waters. Brown et al. (2008) concluded that there was insufficient evidence to link nitrate enrichment to changes in algal cover. Heffernan et al. (2010) argue convincingly that studies by Canfield and Hoyer (1988) and Duarte and Canfield (1990) found no relationships between nutrients and total vegetative biomass in spring runs as would be expected under nutrient limitation scenarios, and that recent surveys of algal biomass have not found any significant linkage of biomass to N or P concentrations (Stevenson et al. 2004; Stevenson et al. 2007). Further, Heffernan et al. (2010) and Liebowitz (2013) report stronger relationships between dissolved oxygen (DO), grazer populations, and algal abundance than with nutrients.
Heffernan et al. (2010) documents a decline in water column DO in Silver Spring and suggests looking to other drivers of algal proliferation in spring systems, including DO control of invertebrate grazers that may result in altered trophic structure in springs to favor algal dominance. Liebowitz (2013) reports a significant negative association between algal and gastropod biomass in Florida springs suggesting top down control of algae by invertebrate grazers, a finding supported by several studies of grazer control of algae in other systems (Hildebrand 2002; Heck and Valentine 2007; Gruner et al. 2008; Baum and Worm 2009; Estes et al. 2011). Further, Liebowitz (2013) also found a significant relationship between dissolved oxygen (DO) and gastropod biomass in a survey of 11 springs, suggesting DO has a significant indirect effect on algal biomass via controlling grazer abundance and/or activity. Under low flow or current velocity conditions, nutrient enrichment and subsequent algal growth may outpace grazer pressure resulting in severe light reductions (Harlin and Thorne-Miller 1981). Alternatively, under similar nutrient enrichment and moderate to high flushing or exchange of water (as in lotic or tidally influenced systems), herbivores have been observed to control epiphytic algal biomass (Neckles et al. 1993. Liebowitz (2013) argues that hysteretic responses of grazer populations to disturbances could be responsible for the over abundance of algae in springs where no clear grazer stress is present. For instance, invasive plant control measures utilizing herbicides and copper compounds are widely employed with known negative impacts on grazer populations (Evans 2008). Such a disturbance could enable algal populations to exceed thresholds for grazer control. This gives rise to a second hypothesis (H2) that grazer control of algae in springs has been altered by DO in some cases and by episodic or unknown exposures to other stressors.
The presence of herbicides or other agrochemicals that may be inhibitory to either algal grazers or macrophytes themselves supports a third hypothesis (H3) which states that a “nitrate cohort” defined as substance[s] associated with the same mechanisms involved in increased nitrate concentration, such as changes to land use (concentrated feed lots for cattle) or ineffective waste treatment (spray fields or failed septic tanks) has an inhibitory or toxic effect on SAV. The widespread use of agro-chemicals such as commercial pesticides, fungicides and herbicides increases poten-tial for these compounds to impact spring ecosystems. Several anthropogenic organic compounds have been detected in springs; however, the low levels observed did not elicit alarm (Phelps et al. 2006; Phelps 2004). Recent increases in consumer use of compounds such as atrazine, a herbicidal fertilizer additive, (Ackerman 2007) and triclosan, an antimicrobial agent, (Fulton et al. 2010) suggest these compounds may have deleterious effects on SAV (and/or grazer population dynamics).
A fourth and least well understood hypothesis (H4) is that nitrate itself has an inhibitory (direct) effect on SAV growth resulting in a shift to algae dominated system. The accepted view of nitrate and ammonia combined with P to create conditions for algal dominance and subsequent shading out of macrophytes (Mulligan et al. 1976) is that of an indirect effect. However, a direct effect of nutrient enrichment has been suggested as a factor in macrophyte disappearance in aquatic systems undergoing enrichment (Genevieve et al. 1997; Farnsworth and Baker 2000). Several authors (Klotzli 1971; Schroder 1979; Boar et al. 1989), have reported correlation of reed bed disintegration and increases in nitrate loading to lakes in England. Decreases in Phragmites australis root and rhizome production was observed in concert with increased nitrate loading (Ulrich and Burton 1985). Ulrich and Burton (1985) also reported that nitrate stimulated growth and overall biomass increased with increased nitrate availability, however, below ground biomass production (roots and rhizomes) did not increase at concentrations up to 6 mg NOx-N L-1. These NOx-N concentrations resulted in significant decreases in below ground to above ground biomass ratios and resulted in an overall decline in health of the reed stands. Nitrate to potassium ratios in surface waters and in tissues are correlated with highest degree of degradation of Phragmites australis beds (Boar et al. 1989), however, the causative mechanism is unknown as is the potential for synergistic effects of increased availability of N and K. Because the aforementioned species is an emergent macrophyte, free of algal shading, it serves as a significant indicator of potential inhibitory effects of NOx-N on plant growth. An in depth review of current literature suggests several authors have observed apparent direct inhibitory effects of NOx-N on SAV in both marine and freshwater environments. The remainder of this paper will synthesize the available literature and investigate potential causal mechanisms for the observed negative effects of elevated NOx-N on SAV.
Evidence of Direct Effects of Nitrate on SAV
Opportunistic luxury consumption of nu-trients is characteristic of SAV and thus accumulation in tissues is anticipated for macrophytes adapted to limited nutrient availability, such as seagrasses and some freshwater SAV (Wetzel 2001). The pre-vailing viewpoint is that most macrophytes acquire nitrogen via roots (Cedergreen and Madsen 2003), however, foliar absorption is also a viable mechanism when sediment sources are not available or abundant (Barko and Smart 1986). When ammonium nitrogen (NH4-N) concentration exceeds 0.1 mg L-1, macrophytes preferentially use NH4-N (Nichols and Keeny 1976). Hence, the dominant form of N utilized by most SAV is NH4-N. However under N limitation ni-trate is also utilized, predominately from the water column. Due to its abundance, NOx-N is the presumed dominant form for N uptake by SAV in Florida springs. Several researchers have made qualitative observations of SAV inhibition closest to spring vents where NOx-N concentrations are highest (Munch et al. 2006; Mattson pers. observation). Similarly, several authors suggest observed declines in macrophytes in other systems was a direct effect of increased NOx-N (Burkholder et al. 1992; Burkholder et al. 1994; Wang et al. 2012), suggesting closer investigation of this phenomenon is warranted.
Burkholder et al. (1992) report that Zostera marina (eelgrass) exhibited highly negative physiological effects (even death) when dosed with 0.05, 0.1 and 0.5 mg L-1 NOx-N. Although a marine species, this plant shows extreme sensitivity to increased nitrate evidenced through loss of carbon storage in roots unrelated to shading by algae. The apparent lack of an inhibition or regulation mechanism of nitrate uptake by eelgrass (Roth and Pregnall 1988) was implicated in the observed disruption of internal nutrient ratios, presumably due to carbon expenditure in amino acid synthesis to reduce intracellular ammonia toxicity. Hierarchical partitioning analysis of water quality parameters found NOx-N exerted the greatest detrimental effect on charophyte occurrence in wetlands of the UK (Lambert and Davy 2011). In situ studies of Chara globularis showed that it was extremely sensitive to nitrate with maximal relative growth rate observed at 0.5 mg NOx-N L-1 and a linear decline in growth with higher concentrations. At 6 mg NOx-N L-1, growth was severely limited, similar to results of no NOx-N treatment (Lambert and Davy 2011). Similarly, biomass accumulation was strongly inhibited by nutrient accumulation (N) in Potamogeton maackianus A. Been (Ni 2001). The most definitive observations of inhibition were in the form of shrinkage of arenchyma tissues and disappearance of starches and chloroplasts observed in increased NOx-N and NH4-N concentration treatments of Vallisneria natans (Wang et al. 2012.). It is important to note that ammonia, whether sourced from high concentrations in water or derived (via intracellular nitrate reduction) from elevated nitrate availability to SAV is the source of toxicity.
The paradigm of nitrogen effects on water clarity often overshadows potential direct effects of excess N on SAV. For instance, Sagrario et al. (2005) reported that high N is not directly inhibitory to Potamogeton pectinatus L., Elodea canadensis and Nymphea sp. at 10 mg L-1 of total N (TN) due to overpowering effects of increased algal shading. However, closer inspection of the results indicates moderate dosing of 4 mg TN L-1 resulted in decreased growth with respect to controls under equal or better water clarity, a noteworthy result that went unmentioned. Further, summer TN levels declined significantly in mesocosms truncating the duration of exposure for macrophytes, which likely confound interpretation of the results by the authors. In a study by Li et al. (2008), NOx-N additions were noted to increase Vallisneria spinulosa biomass over control at 2.5, 5.0, 7.5 mg L-1 concentrations in water column but at 10 mg L-1 growth was not significantly different from control (1 mg L-1) suggesting some inhibition of growth. It is unclear why the authors did not conclude that a NOx-N threshold had been exceeded between 7.5 and 10 mg L-1. This lack of interpretation by some authors is likely due to a strong focus on algal production and subsequent shading, not direct effects of nitrate on SAV (Sturgis and Murray 1997). Further, variability among species with respect to effects of NOx-N appears to be high (Burkholder et al. 1994). This is exemplified by conflicting reports on potential inhibition of macrophyte growth by excessive water column nitrate (Li et al 2005). Best (1980) reported no inhibition of Ceratophyllum demersum at concentrations of up to 105 mg NOx-N L-1 but did observe ammonia toxicity at 45 mg NH4-N L-1. This finding suggests that C. demersum is well suited to luxury uptake of N. Conversely, Lambert and Davy (2011) assert a mean annual concentration limit of 2 mg NOx-N L-1 is necessary to protect charophytes.
Our review of available literature did not find studies of N enrichment with SAV species common in Florida spring systems (Vallisneria americana, Sagittaria kurziana, Najas spp., Potamogeton spp.). However, the potential mechanism of inhibition, which likely varies among species, are discussed here in general terms for SAV and are viewed as potential mechanisms until tested on individual species of interest.
Potential Mechanisms of Inhibition
Nitrate toxicity has been well documented for vertebrate animals (including humans) (Kim-Shapiro et al. 2005) as well as invertebrates (Mattson et al. 2007). However, the potential of NOx-N toxicity or inhibition of SAV is not well understood, nor is it intuitive given our understanding of mechanisms of toxicity for higher or-ganisms. Observations coinciding with elevated inorganic N (NOx-N and or NH4-N) in plants include stunted growth, iron deficiency, amino acid accumulation, oxidative stress and structural tissue damage (Burkholder et al 1992; Smolders et al. 1997; Smolders et al. 2000; van der Heide et al. 2008; Wang et al. 2012). To better determine potential inhibitory mechanisms of NOx-N, a closer look at the process of nitrogen assimilation is necessary.
Assimilatory Nitrate Reduction
Most aquatic plants absorb nitrate, which is then sequentially converted to nitrite and then ammonium by the nitrate reductase system (Salisbury and Ross 1992). In SAV, before nitrate can be utilized by the plants, it must be converted to ammonium by a series of sequential enzyme mediated reactions (Figure 1) involving nitrate reductase and nitrite reductase (Guerrero et al. 1981). This process is termed assimilatory nitrate reduction (ANR) and results in ammonia being incorporated into amino acids. Genetic or environmental factors, such as light, temperature, depth, pH, and location within vegetated patch (edge versus center) (Roth and Pregnall 1988; van der Heide et al. 2008), can modulate this series of biochemical reactions resulting in a high level of variability among species with respect to nitrate reduction processes and rates (Pate 1980; Guerrero et al. 1981). Water temperature can be problematic for SAV by increasing respiration rates and impairing enzyme function (Zimmerman et al. 1989; Lacoul and Freedman 2006; Riis et al. 2012), thus impairing nitrogen assimilation. However, SAV in spring runs generally do not experience thermal stress due to the thermal consistency of groundwater (unless exposed in shallow backwaters).
Uptake of NOx-N is driven primarily by external nitrate concentrations (Marschner 1998) and in aquatic macrophytes, increased water column concentrations of NOx-N results in significant increases in nitrate reductase activity (NRA) (Cedergreen and Madsen 2003). Studies of Zostera marina indicate newer leaves are more active with respect to NRA and rates between individual plants can be variable with a 2-3 fold difference (Roth and Pregnall 1988). It has been suggested that differences between root and shoot NRA depends upon uptake rates of individual species (Gojon et al. 1994) and that location of nitrate reduction (root or shoot) is also species specific (Cedergreen and Madsen 2003). From an energetic standpoint, photosynthetic tissues would be a more advantageous location for NRA to occur due to a lack of need to transport NOx-N to the roots (Raven 1985; Schjoerring et al. 2002) and this appears to be the case for SAV (Roth and Pregnall 1988). Once uptake has occurred, there are physiological deterrents to storing nitrate freely within the cytoplasm due to osmotic stress (Salisbury and Ross 1992). Therefore nitrate is stored in vacuoles or rapid conversion to ammonia occurs within the cytoplasm (Heimer and Filner 1971; Wagner 1979; Granstedt and Huffaker 1981). These storage vacuoles are not, however, without their limitations. Hydration of vacuolar materials such as nitrate or associated cations (K, Na) can require significant water, which ultimately limits the concentration that can be stored intracellularly (Raven and Smith 1976). Regardless of the storage method of nitrate, the reduction of nitrate and subsequent production of ammonia requires plants to avoid toxicity by allocating carbon and energy to protein (amino acid) synthesis to alleviate ammonia buildup (Salisbury and Ross 1992). Under normal exposure to NOx-N, ANR uses approximately 25% of the reductant energy produced by photosynthesis and root/shoot respiration (Crawford 1995).
![Figure 1. Conceptual model of nitrate overload hypothesis. Uptake of nitrate is unregulated at the cellular level and presence of nitrate induces nitrate reduction to ammonia. Buildup of ammonia should be a negative feedback[-] for nitrate reduction enzymes; however this process appears not to function in some species. Ammonia can be toxic to plants and therefore is alleviated via amino acid and consequent protein synthesis, which requires energetic inputs from plant carbohydrate stores. Buildup of free amino acids and depletion of root carbohydrate stores are potential diagnostics of nitrate overload in SAV.](/uploads/vol8/osborne/Image3510091.JPG "Figure 1. Conceptual model of nitrate overload hypothesis. Uptake of nitrate is unregulated at the cellular level and presence of nitrate induces nitrate reduction to ammonia. Buildup of ammonia should be a negative feedback[-] for nitrate reduction enzymes; however this process appears not to function in some species. Ammonia can be toxic to plants and therefore is alleviated via amino acid and consequent protein synthesis, which requires energetic inputs from plant carbohydrate stores. Buildup of free amino acids and depletion of root carbohydrate stores are potential diagnostics of nitrate overload in SAV.")
Figure 1. Conceptual model of nitrate overload hypothesis. Uptake of nitrate is unregulated at the cellular level and presence of nitrate induces nitrate reduction to ammonia. Buildup of ammonia should be a negative feedback[-] for nitrate reduction enzymes; however this process appears not to function in some species. Ammonia can be toxic to plants and therefore is alleviated via amino acid and consequent protein synthesis, which requires energetic inputs from plant carbohydrate stores. Buildup of free amino acids and depletion of root carbohydrate stores are potential diagnostics of nitrate overload in SAV.
Closer inspection of the biochemical pathways for ANR reveals some significant differences between SAV and filamentous macroalgae, the two competing primary producers in many springs. Assimilatory nitrate reductase activity in green algae and higher plants is dependant upon NAD(P)H for reducing power (Figure 2A). Further, the negative feedback inhibitor of the nitrate reductase enzyme in some species is nitrite, which competitively binds with nitrate reductase. This is not the case for all species as reported by Roth and Pregnall (1988) who documented the inability of Zostera marina to “turn off” or regulate nitrate reductase, a very critical observation with respect to the potential for some SAV to moderate this enzyme as it establishes evidence that some species may not have the capacity to regulate NRA. Cyanobacteria, on the other hand, cannot utilize reduced pyridine nucleotides as do green algae and higher plants. The alternative electron donor for algal nitrate reductase (Figure 2B) is ferredoxin (Guerrero et al. 1981). This reaction appears to give cyanobacteria a slight energetic advantage as the ΔG of the reaction is 4.6 Kcal greater per mole for ferredoxin mediated reduction versus NAD(P)H. The second reduction reaction, nitrite reduction to ammonium, is very similar in all photosynthetic organisms and utilizes ferredoxin as the electron donor specifically.
![Figure 2. Assimilatory nitrate reduction in [A] SAV shoot and [B] cyanobacterial cell. Note cyanobacteria cannot utilize NAD(P)H as an electron donor in the reduction of nitrate to nitrite and thus rely solely upon ferrodoxin. Also note a slight energetic advantage [ΔG’] exists for cyanobacteria in the reduction of nitrate to nitrite.](/uploads/vol8/osborne/Image3510092.PNG "Figure 2. Assimilatory nitrate reduction in [A] SAV shoot and [B] cyanobacterial cell. Note cyanobacteria cannot utilize NAD(P)H as an electron donor in the reduction of nitrate to nitrite and thus rely solely upon ferrodoxin. Also note a slight energetic advantage [ΔG’] exists for cyanobacteria in the reduction of nitrate to nitrite.")
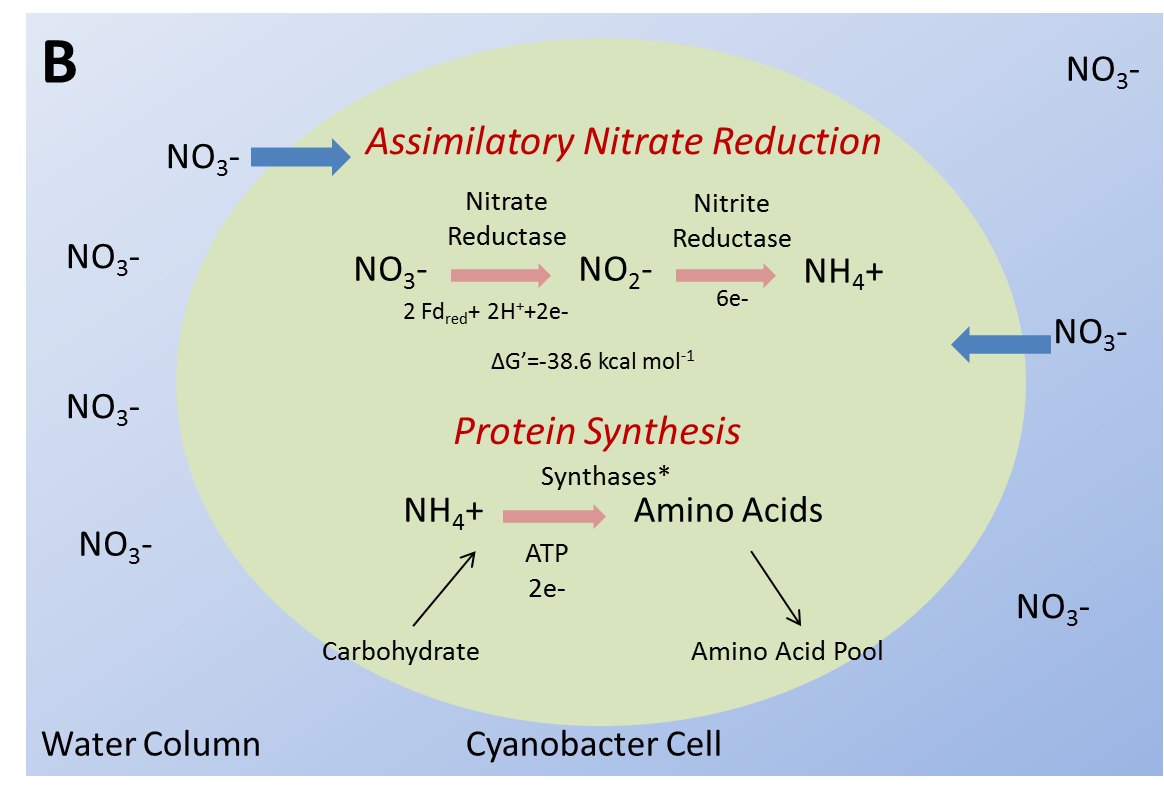
Figure 2. Assimilatory nitrate reduction in [A] SAV shoot and [B] cyanobacterial cell. Note cyanobacteria cannot utilize NAD(P)H as an electron donor in the reduction of nitrate to nitrite and thus rely solely upon ferrodoxin. Also note a slight energetic advantage [ΔG’] exists for cyanobacteria in the reduction of nitrate to nitrite.
Ferredoxin requires iron in its structural complex, thus increased iron concentration in springs may also provide cyanobacteria a competitive advantage over green algae and SAV. This is due to the inability of the latter organisms to utilize ferredoxin in nitrate reduction. Smolders et al. (1997) report iron deficiency in SAV exposed to higher levels of NOx-N, presumably due to the need for ferredoxin in nitrite reduction. Because NAD(P)H also serves as reducing power for many other metabolic reactions, utilization of NAD(P)H for nitrate reductase results in a decrease of is use in other metabolic reactions and potential buildup of other metabolites within cells, which may reduce plant growth and metabolic processes (Lea and Miflin 1979). Of greater concern, the accumulation of ammonia, the end product of ANR, can be extremely detrimental to photosynthetic organisms. Excessive intercellular ammonia repre-sents a significant source of toxicity for SAV through inhibition of respiration, metabolic activities, and compromising of cell membrane integrity. This toxicity may be realized when excess nitrate availability is combined with the possibility of poorly regulated ANR, resulting in ammonia buildup in tissues. ().
Ammonia Toxicity
Ammonia toxicity is well documented in terrestrial plants (Salisbury and Ross 1992) as well as in seagrasses and other hydrophytic vegetation (Katwijik et al. 1997; Hemminga and Duarte 2000). In most plants, excessive ammonia is toxic due to inhibition of respiration, photosynthesis and other metabolic processes. Ammonia toxicity is often indicated by a decrease in soluble sugars in the cytoplasm (Cramer and Lewis 1993) or excessive tissue con-centrations of ammonium that exceed the plants’ ability to incorporate into amino acids (Meher and Mohr 1989). Inhibitory effects of high ammonium on SAV have been documented (Best 1980; Smolders et al. 1996) and implicated in succession of freshwater SAV communities (Schuurke et al. 1986; Brouwer et al. 1997; Clarke and Baldwin 2002). Excessive ammonium can inhibit photosynthesis (Cao et al. 2004) resulting in diminished photosynthate (soluble sugar) production and lead to necrosis in some macrophytes (Smolders et al. 1996). To alleviate NH4-N stress, plants must convert the free ammonium to amino acids via biosynthesis (Figures 1 and2). This process has an energetic cost, requiring carbon and energy inputs from the plant (i.e. soluble sugars, photosynthate). Lambert and Davy (2011) invoke energetic expenditure in regulating ammonia internally as a likely cause of growth decline in Chara sp. exposed to NOx-N in excess of 2 mg L-1. Water column ammonia concentrations >1 mgL-1 resulted in decreased soluble sugar content in Potamogeton crispus and increased soluble amino acids (Cao et al. 2004). Interestingly, in a study by Cao et al. (2004), responses of amino acids and soluble sugar indicators of ammonia stress were dependant upon duration of exposure. Further, activity of ascorbate peroxidase and superoxide dismutase (both anti-oxidant enzymes) were highest at 1mg L-1 ammonium and decreased significantly as ammonium increased (Cao et al. 2004). Other factors are known to influence the degree of ammonia toxicity in plant tissues. For instance, Netten et al. (2013) reported that light and temperature also had significant effects on toxicity of ammonia in Elodea Canadensis. While temperature and water column ammonia concentration are not issues for the spring ecosystems discussed here (Munch et al. 2006), the availability of NOx-N can provide excessive nitrogen to SAV.
The energetic demand of reducing the resulting ammonia toxicity, in concert with unregulated ANR, could represent a very significant stress on SAV (Smolders et al. 2000; Wang et al. 2012).
Amino-Acid Synthesis
Ammonia is incorporated into α-amino-acids by way of one or both known pathways (Figure 2), the glutamate dehydrogenase and the glutamate synthetase-glutamate synthase pathway (Guerrero et al. 1981). Buildup of free amino acids in tissues is considered an indication of “nitrogen overload” or impending toxicity due to excessive nitrogen availability (Smolders et al. 1996; Smolders et al. 2000; Wang et al. 2012). Specific types of amino acids that accumulate in tissues (for instance arginine, glutamine, asparagines) are dependant on the stresses involved (toxicity, mineral deficiency, grazer pressure) and the species of SAV (Rabe and Lovatt 1986; Rabe 1990; Marschner 1998; Smolders et al. 2000). Significant evidence of the nitrogen overload hypothesis is presented by Wang et al. (2012) who reported reduction of arenchyma tissue, chloroplasts and starch grains in tissues of Vallisneria natans exposed to increased nitrate and ammonia levels. The authors contend that loss of structures and starch content is related to photosynthate required to reduce nitrate to ammonia and further sequester toxic ammonia in amino acids, a process that requires significant energy expenditure by plants. Due to the high energetic demand, NOx-N overload may perhaps lead to susceptibility to pathogens. For example, Zostera marina, as well as some other angiosperms, are known to decrease production of antimicrobial compounds such as phenolics during times of increased protein synthesis associated with N enrichment (Buchsbaum et al. 1990).
Summary of Mechanisms of Inhibition
Review of the current literature concerning NOx-N effects on SAV is compelling in that the process of ANR is highly variable among species and the potential for unregulated uptake, an adaptation ostensibly stemming from luxury uptake, could induce the “nitrogen overload” condition (Smolders et al. 1996; Smolders et al 2000; Boedeltje et al. 2005; Wang et al. 2012). The resulting accumulation of ammonia, the end product of ANR, can itself be a significant stressor to plants or, by necessitating protein synthesis to alleviate ammonia stress, can cause depletion of SAV carbohydrate stores (Guerrero et al. 1981; Wang et al. 2012). In other aquatic ecosystems (ponds, lakes) that are finite in the overall mass of N, potential NOx-N toxicity may be reduced based upon density of SAV (van der Heide et al. 2010). However, under the unique lotic conditions of springs (increasing nitrate concentrations and constant exposure due to high flow conditions), the potential for increased biomass of SAV to ameliorate potential toxicity is not anticipated. Determining these direct effects of NOx-N on SAV native to Florida springs will be of primary importance to directing management effort with respect to springs restoration.
Ecological Implications
Globally, many aquatic ecosystems have been altered, some seemingly irrevocably, by the anthropogenic addition of excessive nutrients (N and P). For example, in both temperate and tropical lakes undergoing nutrient enrichment, catastrophic shifts from macrophyte to phytoplankton dominance have been observed with regularity in the last half century. In Florida, significant effort has been invested in ameliorating these catastrophic shifts on large lakes such as Apopka (Dunne et al. 2012) or Okeechobee (James et al. 2011; Harwell and Sharfstein 2009). This shift in primary productivity has resounding effects throughout the food web. Further, habitat loss and susceptibility to altered environmental conditions (for example: hypoxia, shifts in pH) can have detrimental effects on established flora and fauna. This shift in primary productivity may also impart significant changes to ecosystem services such as biogeochemical cycling of nutrients and habitat quality for fauna. Similarly, Florida’s springs systems, which have immense ecological, cultural and economic value to the state have undergone significant ecological degradation in recent decades. Therefore, concern exists for determining the relationship between these changes and the observed increase in NOx-N in springs. Of primary concern is elucidating the role nitrate enrichment has had (whether direct or indirect via synergistic interactions with other stressors) in the observed decline of these systems.
Research Needs
Research is warranted to investigate if SAV native to Florida springs are experiencing any inhibitory effects due to elevated NOx-N concentrations by one or more of the proposed mechanisms: 1) unregulated NOx-N uptake and reduction, 2) ammonia toxicity from excess accumulation in vivo, and 3) carbohydrate depletion from intercellular or root storages. Of primary interest would be a synoptic sampling of SAV from several springs along gradients of elevated NOx-N, as well as sampling springs without significant increases of NOx-N, to determine if gradients of inhibition or energetic stress is observed. Species of interest are Vallisneria americana, Sagittaria kurziana,Najas sp., Potamogeton sp., Ceratophyllum spp., Hydrilla sp and Chara spp. Biometrics such as root and shoot biomass and calculation of root:shoot mass ratios may be the most informative preliminary measurement approach. If these preliminary results suggest significant impact to growth from elevated NOx-N, then more intensive studies may be indicated. These studies might include, but are not limited to, determination of direct use of NOx-N by SAV, toxicity thresholds for NOx-N (via NH4-N) in tissues, and potential synergistic effects of NOx-N, K, and Fe. With consideration of the significant changes occurring in Florida Spring ecosystems, other potential contributors to algal proliferation (indirect effect on SAV health) such as declining DO and multiple stressor hypotheses also warrant investigation.
Discussion with Reviewers
1. Reviewer 2: Could the authors add a comment on the molecular basis for ammonia toxicity in animals (Monfort et al. 2002)?
While this contribution deals specifically with submerged aquatic vegetation, the authors realize that internal physiological nitrate/nitrite (NOx-N) reduction to ammonia by invertebrates in the same ecosystem could bring about a loss of top down control of algae (and concomitant decline in SAV) if algal grazing organisms were negatively affected by increased NOx-N. Monfort et al. (2002) present a molecular basis for ammonia toxicity in which activation of N-methyl-D-aspartate (NMDA) is indicated during excessive ammonia exposure. Strong evidence is presented supporting the assertions that NMDA receptors are responsible for the following effects (from Monfort et al. 2002): exhaustion of ATP levels in the brain; dephosphorylation and activation of Na+/K+-ATPase in brain; impairment of mitochondrial function and calcium homeostasis which decreases ATP synthesis; reduction of glutamine synthetase activity which reduces ammonia elimination in the brain. Combined these effects reduce brain activity and neuronal degeneration and eventual mortality. The process outlined by Monfort et al. (2002) has application to this work in that it could be an additional stressor on the grazer community resulting in unchecked algal growth (a common observation in springs with impaired SAV). Moreover, the process described alters ATP availability and production in the brain tissue of invertebrates and is similar in that respect to the process proposed in our manuscript which invokes the depletion of photosynthate stores in SAV tissues during amino acid synthesis which is required to reduce intracellular levels of ammonia resulting from unchecked NOx-N reduction.
Monfort P, Kosenko E, Erceg S, Canales J-J, Felipo V (2002) Molecular mechanism of acute ammonia toxicity: role of NMDA receptors. Neurochemistry International 41(2-3): 95-102)
2. Reviewer 2: The authors also ought to consider a molecular basis for ammonia toxicity, resulting from the Maillard and the Strecker reactions.
The authors did not describe the molecular basis for ammonia toxicity in the paper, rather for brevity sake, simply referred to its occurrence only. The actual mode of toxicity from accumulating ammonia in the cells of SAV may be sourced from intracellular production of physiologically disrupting compounds produced by common sequential reactions of amino acids sourced from large quantities of free ammonia building up in plant cells. Hence, Maillard and Strecker reactions have relevance to this discussion. First, the Maillard reaction is a chemical reaction that produces glycosamine from amino groups of amino acids reacting with carbonyl groups of reducing sugars within the cell. Glycosamine is an unstable product and thus undergoes further reactions (Amadori series rearrangement) which yields ketosamines. These compounds, after undergoing further dehydration and de-amination, produce di-carbonyls. The Strecker reaction is involved when the di-carbonyl products react further with readily available amines to produce acrylamide, a known carcinogen and cellular disruptor. This is especially the case when asparagine is present (a dominant amino acid observed in SAV under high nitrate conditions by Wang et al. 2012). Thus, while the authors observe increased nitrate/ nitrate in the spring ecosystems and focus on the role this NOx-N may have on SAV in the systems, the actual process is a bit more complicated in that NOx-N reduction to ammonia is not in itself phyto-toxic, rather, the buildup of ammonia and resulting reactions such as the Maillard and Strecker reactions, can produce toxicity at the cellular level. We hypothesize here that SAV, in an effort to alleviate toxicity of lesser regulated ammonia production, utilizes stored energy from photosynthate to produce amino-acids. The authors recognize that under specific conditions, either mechanism could produce the observed stunting (and possibly loss) of SAV observed in Florida Springs.
Acknowledgements
The authors would like to thank John Hendrickson, Steve Miller, and Andy Canion for their thoughtful reviews and Jennifer Hornsby for her editorial assistance in preparation of this manuscript. This work was supported by the St. Johns River Water Management District.
References
Ackerman, F. 2007. The economics of atrazine. International Journal of Occupational and Environmental Health. 13:437-445.
Albertin, A.R. 2009. Nutrient dynamics in Florida springs and relationships to algal blooms. Dissertation, University of Florida.
Albertin, A.R., J.O. Sickman, A. Pinowska, R.J. Stevenson. 2012. Identification of nitrogen sources and transformations within karst springs using isotope tracers of nitrogen. Biogeochemistry 108: 219-232.
Barker, T., K. Hatton, M. O’Connor, L. Connor, B. Moss. 2008. Effects of nitrate load on submerged plant biomass and species richness: results of a mesocosm experiment. Fundamental and Applied Limnology 173: 89-100.
Barko J.W. and R. M. Smart. 1986. Sediment-related mechanisms of growth limitation in submersed macrophytes Ecology 67: 1328-1340.
Baron, J.S., E.K. Hall, B.T. Nolan, J.C. Finlay, E.S. Bernhardt, J.A. Harrison, F. Chan, E.W. Boyer. 2013. The interactive effects of excess reactive nitrogen and climate change on aquatic ecosystems and water resources of the United States. Biogeochemistry 114: 71-92.
Baum, J. K., and B. Worm. 2009. Cascading top-down effects of changing oceanic predator abundances. Journal of Animal Ecology 78:699–714.
Bergstrom, A.K., and M. Jansson. 2006. Atmospheric nitrogen deposition has caused nitrogen enrichment and eutrophication of lakes in the northern hemisphere. Global Change Biology 12: 635-643.
van den Berg, M.S., M. sheffer, E. van Nes, H. Coops. 1999. Dynamics and stability of Chara sp. and Potamogeton pectinaus in a shallow lake changing in eutrophication level. Hydrobiologia 408: 335-342.
Best, E.P.H. 1980. Effects of nitrogen on the growth and nitrogenous compounds of Ceratophyllum demersum. Aquatic Botany 8: 197-206.
Boar, R.R., C.E. Crook, B. Moss. 1989. Regression of Phragmites ausitralis reed swamps and recent changes if water chemistry in the Norfolk Broadland, England. Aquatic Botany 35: 41-55.
Boedeltje, G., A.J. Alfons, J.P. Smolders, J.G.M. Roelofs. 2005. Combined effects of water column nitrate enrichment, sediment type and irradiance on growth and foliar nutrient concentrations of Potamogeton alpines. Freshwater Biology 50: 1537-1547.
Borum, J. 1985. Development of epiphytic communities on eelgrass (Zostera marina) along a nutrient gradient in a Danish estuary. Marine Biology 87: 211-218.
Brouwer, E., R. Bobbink, F. Meeuwsen, J.G.M. Roelofs. 1997. Recovery from acidification in aquatic mesocosms after reducing ammonium and sulfate deposition. Aquatic Botany 56: 119-130.
Brown, M.T., K. Chinner-Reiss, M.J. Cohen, J.M. Evans, K.R. Reddy, P.W. Inglett, K.S. Inglett, T. K. Frazer, C.A Jacoby, E.J. Philips, R. L. Knight, S. K. Notestein, K.A. McKee. 2008. Summary and synthesis of the available literature on the effects of nutrients on springs organisms and systems. Final Report. Florida Department of Environmental Protection, Tallahassee FL.
Buchsbaum, R.N., F.T. Short, D.P. Cheney. 1990. Phenolic-nitrogen interactions in eelgrass (Zostera marina L.): possible implications of disease resistance. Aquatic Botany 37: 291-297.
Burkholder, J.M., K.M. Mason, H.B. Glasgow, Jr. 1992. Water column nitrate enrichment promotes decline of eelgrass Zostera marina: evidence from seasonal mesocosm experiments. Marine Ecological Progress Series 81: 163-178.
Burkholder, J.M., H.B. Glasgow, J.E. Cooke. 1994. Comparative effects of water column nitrate enrichment on eelgrass Zostera marina, shoal grass Halodule wrightii and widgeongrass Ruppia maritime. Marine Ecological Progress Series 105:121-138.
Canfield, H.E., and M.V. Hoyer. 1988. Influence of nutrient enrichment and light availability on the abundance of aquatic macrophytes in Florida streams. Canadian Journal of Fisheries and Aquatic Sciences 45: 1467-1472.
Cao, T., L. Ni, P. Xie. 2004. Acute biochemical responses of a submersed macrophyte, Potamogeton crispus L., to high ammonium in an aquarium experiment. Journal of Freshwater Ecology 19: 279-284.
Cedergreen, N. and T.V. Madsen. 2003. Nitrate reductase activity in roots and shoots of aquatic macrophytes. Aquatic Botany 76: 203-212.
Clarke, E. and H.A. Baldwin. 2002. Responses of wetland plants to ammonia and water level. Ecological Engineering 18: 257-264.
Cohen, M.J., S. Lamsal and L.V. Korhnak. 2007. Sources, Transport and Transformations of Nitrate-N in the Florida Environment. St. Johns River Water Management District SJ2007-SP10. 125 pp. Palatka, FL.
Cowell, B.C. and P.S. Botts. 1994. Factors influnceing the distribution, abundance, and growth of Lyngbya wollei in Central Florida. Aquatic Botany 49: 1-17.
Cowell, B.C. and C.J. Dawes. 2004. Growth and nitrate nitrogen uptake by the cyanobacterium Lyngbya wollei. Journal of Aquatic Plant Management. 42: 69-71.
Cowell, B.C., C.J. Davis. 2008. Seasonal comparisons of the phytoplankton and decline in water transparency in spring-fed rainbow River, Florida. Journal of Freshwater Ecology 23: 169-177.
Cramer, M.D. , O.A. M. Lewis. 1993. The influence of NO3 and NH4 nutrition on carbon and nitrogen partitioning characteristics of wheat (Triticum aestivum) and maize (Zea maize) plants. Plant and Soil 154: 289-300.
Crawford, N.M. 1995. Nitrate: nutrient signal for plant growth. The Plant Cell 7:859-868.
Duarte, C.M. and D.E. Canfield, Jr. 1990. Macrophyte standing crop and primary productivity in some Florida spring-runs. Water Resources Bulletin 26(6): 927-934.
Dunne, E.J., M.F Coveney, E.R. Marzolf, V.R. Hogue, R. Conrow, R. Naleway, E.F. Lowe, L.E. Battoe. 2012. Efficacy of a large-scale constrcutied wetland to remove phosphorus and suspended solids from Lake Apopka, Florida. Ecological Engeineering 42: 90-100.
Dzialowski, A.R., S.H. Wang, N.C. Lim, W.W. Spotts. 2005. Nutrient limitation of phytoplankton growth in central plains reservoirs, USA. Journal of Plankton Research 27: 587-595.
Elser, J.J., M.E.S. Bracken, E.E. Cleland, D.S. Gruner, W.S. Harpole, H. Hillebrand, J.T. Ngai, E.W. Seabloom, J.B. Shurin, J.E. Smith. 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine , and terrestrial systems. Ecological Letters 10: 1135-1142.
Evans, J. 2008. Ecosystem implications of invasive aquatic plants and aquatic plant control in Florida Springs. P 249-270 In: M.T. Brown, Ed. Summary and synthesis of available literature on the effects of nutrients on springs organisms and systems. Florida Department of Environmental Protection. Tallahassee, FL.
Estes, J. a, J. Terborgh, J. S. Brashares, M. E. Power, J. Berger, W. J. Bond, S. R. Carpenter, T.E. Essington, R. D. Holt, J. B. C. Jackson, R. J. Marquis, L. Oksanen, T. Oksanen, R. T. Paine, E. K. Pikitch, W. J. Ripple, S. a Sandin, M. Scheffer, T. W. Schoener, J. B. Shurin, A. R. E. Sinclair, M. E. Soulé, R. Virtanen, and D. Wardle. 2011. Trophic downgrading of planet Earth. Science 333:301–6.
Farnswoth, L., and L.A. Baker. 2000. Conceptual model of aquatic plant decay and ammonium toxicity for shallow lakes. Journal of Environmental Engineering. 126: 199-207.
Frazer, T.K., S.K. Notestein, W.E. Pine. 2006. Changes in the physical, chemical and vegetative characteristics of the Homosassa, Chassahowitzka and Weeki Wachee rivers. Final Report. Southwest Florida Water Mangement District, Brooksville, FL.
Fulton, B.A., R. A. Brain, S. Usenko, J.A. Back, B.W. Brooks. 2010. Exploring Lemna gibba thresholds to nutrient and chemical stressors: differential effects of triclosan on internal stoichiometry and nitrate uptake across a nitrogen: phosphorus gradient. Environmental Toxicology and Chemistry 29:2363-2370.
Genevieve, M.C, H.C Duthie, W.D. Taylor. 1997. Models of aquatic plant productivity: a review of the factors that influence growth. Aquatic Botany 59: 195-215.
Gojon, A. , C. Plassard, C. Bussi. 1994. Root/shoot distribution of NO3- assimilation in herbaceous and woody species. In: Roy, J. and E. Garnier (Eds.) A Whole Plant Perspective on Carbon – Nitrogen Interactions. SPB Academic Publishing, Hague, pp 131-148.
Granstedt, R.C., R.C. Huffaker. 1981. Vacuolar nitrate and the isolation of vacuoles for localization and transport studies. P. 569-577 In Lyons, J.M., R.C. Valentine, D.A. Phillips, D.W. Rains, R.C. Huffaker (eds.). 1981 Genetic Engineering of symbiotic nitrogen fixation and conservation of fixed nitrogen. Plenum Press, New York.
Gruner, D. S., J. E. Smith, E. W. Seabloom, J. T. Ngai, S. Sandin, H. Hillebrand, W. S. Harpole, J. J. Elser, E. E. Cleland, M. E. S. Bracken, E. T. Borer, and B. Bolker. 2008. A cross system synthesis of consumer and nutrient resource control on producer biomass. Ecology Letters 11:740–755.
Guerrero, M.G., J.M. Vega, M. Losada. 1981. The assimilatory nitrate reducing system and its regulation. Annual Reviews of Plant Physiology 32: 169-204.
Harlin, M.M., and B. Thorne-Miller. 1981. Nutrient enrichment of seagrass beds in a Rhode Island coastal lagoon. Marine Biology. 65: 221-229.
Harwell, M. C., B. Sharfstein. 2009. Submerged aquatic vegetation and bulrush in Lake Okeechobee as indicators of greater Everglades ecosystem restoration. Ecological Indicators 9: S46-S55.
Heck, K. L. J., and J. F. Valentine. 2007. The primacy of top-down effects in shallow benthic ecosystems. Estuaries and Coasts 30:371–381.
Heffernan, J. B., D. M. Liebowitz, T. K. Frazer, J. M. Evans,and M. J. Cohen. 2010. Algal blooms and the nitrogen enrichment hypothesis in Florida springs: evidence, alternatives, and adaptive management. Ecological Applications 20:816–829.
van der Heide, T, A.J.P. Smolders, B.G. A. Rijkens, E.H. van Nes, M.M. van Katwijk, J.G.M. Roelofs. 2008. Toxicity of reduced nitrogen in Eelgrass (Zostera marins) is highly dependent on shoot density and pH. Oecologia 158: 411-419.
van der Heide, T., E.H. van Nes, M.M. van Ktawijk, M. Scheffer, A.J. Hendriks, A.J.P. Smolders. 2010. Alternative stable states driven by density-dependent toxicity. Ecosystems 13: 841-850.
Heimer, Y., and Filner, P. 1971. Regulation of the nitrate assimilation pathway in cultured tobacco cells. III. The nitrate uptake system, Biochim. Biophys. Acta, 230:362.
Hemminga, M.A., and C.M. Duarte. 2000. Seagrass Ecology. Cambridge University Press, Cambridge UK. 298 pp.
Hillebrand, H. 2002. Top-down versus bottom-up control of autotrophic biomass—a meta analysis on experiments with periphyton. Journal of the North American Benthological Society 21:349–369.
James,C., J. Fisher, B. Moss. 2003. Nitrogen driven lakes: the Shropshire and Cheshire Meres. Archif fur Hydrobiologia 164: 249-266.
James, C., J. Fisher, V. Russell, S. Collings, B. Moss. 2005. Nitrate avilabliity and hydrophyte species richness in shallow lakes. Freshwater Biology 50:1049-1063.
James, R.T., W.S. Gardner, M.J. McCarthy, S.A. Carini. 2011. Nitrogen dynamics in Lake Okeechobee: forms, functions, and changes. Hydrobiologia 669: 199-212.
Jones, G.W., S.B. Upchurch, K.M. Champion. 1996. Origin of nitrate in ground water discharging from Rainbow Springs, Marion County, Florida. Southwest Florida Water Management District, Brooksville, FL.
Katwijik, M.M.V., L.H.T. Vergeer, G.H.W. Schmitz, J.G.M. Reolofs. 1997. Ammonium toxicity in eelgrass Zostera marina. Marine Ecology Progress Series 157: 159-173.
Katz, B.G. 2004. Sources of nitrate contamination and age of water in large karstic springs in Florida. Environmental Geology 46:689-706.
Kim-Shapiro, D.B., M.T. Gladwin, R.P. Patel, N. Hogg. 2005. The reaction between nitrite and hemoglobin: the role of nitrite in hemoglobin mediated hypoxic vasodilation. Journal of Inorganic Biochemistry 99: 237-246.
Klotzli, F. 1971. Biogenous influence on aquatic macrophytes especially Phragmites communus. Hydrobiologia 12: 107-111.
Lacoul, P., and B. Freedman. 2006. Environmental influences on aquatic plants in freshwater ecosystems. Environmental Reviews 14: 89-136.
Lambert, S. J., and A.J. Davy. 2011. Water quality as a threat to aquatic plants: discriminating between the effects of nitrate, phosphate, boron and heavy metals on Charophytes. New Phytologist 189: 1051-1059.
Lea, P.J., and B.J. Miflin. 1979. Photosyntheitc ammonia assimilation. In: Encylopedia of Plant Physiology. Ed. M. Gibbs, E. Latzko. Springer, Berlin 578pp.
Li, W., Z. Zhang, E. Jeppesen. 2008. The response of Vallisneria spinulosa (Hydrocharitaceae) to different loadings of ammonia and nitrate at moderate phosphorus concentration: a mesocosm approach. Freshwater Biology 53: 2321-2330.
Liebowitz, D.M. 2013. Environmentally mediated consumer control of algae proliferation in Florida springs. Doctoral Dissertation, University of Florida, Gainesville, FL.
Liebowitz, D.M., M.J. Cohen, J.B. Heffernan, T.K. Frazer, L.V. Korhnak. (In review) Environmentally mediated consumer control of algae proliferation in Florida springs.
Maberly, S.C., L. King, M.M. Dent, R.I Jones, C.E. Gibson. 2002. Nutrient limitation of phytoplankton and periphyton growth in upland lakes. Freshwater Biology 47: 2136-2152.
Maddox, G.L., J.M. Lloyd, T.M. Scott, B. Upchurch, R. Copeland. 1992. Florida’s groundwater quality monitoring program, background geochemistry. Florida Geological Survey, Tallahassee, FL.
Marschner, H. 1998. Mineral Nutrition of Higher Plants. Academic Press, London, pp. 232-242.
Mattson, R.A., M. Lehmensiek, E.F. Lowe. 2007. Nitrate toxicity in Florida springs and spring-run streams: a review of the literature and its implications. Professional Paper SJ2007-PP1. St. Johns River Water Management District, Palatka, FL.
Mattson, R.A. Personal Observation. July 19, 2013
Mehrer, I., and H. Mohr. 1989. Ammonium toxicity: description of the syndrome in Sinapis alba and search for its causation. Physiologia Plantarum 77: 54554.
Moss, B. 1990. Engineering and biological approaches to restoration from eutrophication of shallow lakes in which aquatic plant communities are important components. Hydrobiologia 200: 367-377.
Mulligan, H.F., A. Baranowsky, R. Johnson. 1976. Nitrogen and phosphorus fertilization of aquatic vascular plants and algae in replicated ponds. Initial responses to fertilization. Hydrobiologia 48: 109-116.
Munch, D.A., D.J. Toth. C. Huang, J.B. Davis, C.M. Fortich, W.L. Osburn, E.J. Phlips, E.L Quinlan, M.S. Allen, M.J. Woods, P.Cooney, R.L Knight, R.A. Clarke, S.L. Knight. 2006. Fifty year retrospective study of the ecology of Silver springs, Florida. Special Publication SJ2007-SPA. St. John’s River Water Management District, Palatka.
Neckles, H.A., R.L. Wetzel, R.J. Orth. 1993. Effects of nutrient enrichment and grazing on epiphyte-macrophyte (Zostera marina L.) dynamics. Oecologia 93: 285-295.
Netten, J.J.C., T. van der Heide, A.J.P Smolders. 2013. Interactive effects of pH, temperature, and light during ammonia toxicity events in Elodea Canadensis. Chemistry and Ecology DOI:10.1080/02757540.2013.769971
Ni, L. 2001. Effects of water column nutrient enrichment on the growth of Potamogeton maackianus A. Been. Journal of Aquatic Plant Management 39: 83-87.
Nichols, D.S. and D.R. Keeney. 1976. Nitrogen nutrition of Microphyllum spicatum: uptake and translocation of 15N by ramets and roots. Freshwater Biology 6: 145-154.
Odum, H.T. 1957. Trophic structure and productivity of Silver Springs, Florida. Ecological Monographs 27(1): 55-112.
Pate, J.S. 1980. Transport and partitioning of nitrogenous solutes. Annual Review of Plant Physiology 31: 313-340.
Phelps, G. G. 2004. Chemistry of Ground Water in the Silver Springs Basin, Florida, with an Emphasis on Nitrate. U.S. Geological Survey Scientific Investigations Report 2004-5144. vi, 54 pp.
Phelps, G.G., S.J. walsh, R.M. Gerwig, W.B. Tate. 2006. Characterization of the hydrology, water chemistry, and aquatic communities of selected springs in the St. Johns river Water Management District, Florida, 2004. Open-file report 2006-1107. USGS Reston, VA.
Pinowska, A., R.J. Stevenson, J.O. Sickman, M. Anderson. 2007. Integrated interpretation of survey for determining nutrient thresholds for macroalgae in Floirda springs: macroalgal relationships to water, sediment, nutrients, diatom indicators, and land use. Florida Department of environmental Protection, Tallhalssee, FL.
Porter, E.M., W. D. Bowman, C.M. lark, J.E. Compton, L.H. Pardo, J.L Soong. 2013. Interactive effects of anthropogenic nitrogen enrichment and clomate change on terrestrial and aquatic biodiversity. Biogeochemistry 114: 93-12.
Quinlan, E.L., E.J. Phlips, K.A. Donnelly, C.H. Jett, P. Sleszynski, S. Keller. 2008. Primary producers and nutrient loading in Silver Springs, FL, USA. Aquatic Botany 88: 247-255.
Rabe, E. 1990. Stress physiology: the functional significance of the accumulation of nitrogen containing compounds. Journal of Horticultural Science 65: 231-243.
Rabe, E., and C.J. Lovatt. 1986. Increased arginine biosynthesis during P deficiency. Plant Physiology 81: 774-779.
Raven, J.A. 1985. Regulation of pH and generation of osmolarity in vascular plants: a cost-benefit analysis in relation to efficiency of use of energy, nitrogen, and water. New Phytologist 101: 25-77.
Raven, J.A., F.A. Smith. 1976. Nitrogen assimilation and transport in vascular land plants in relation to intracellular pH regulation. The New Phytologist 76: 415-431.
Reddy, K.R and R.L. DeLaune. 2008. Biogeochemistry of Wetlands: Science and Applications. CRC Press Boca Raton. 774 pp.
Riis, T., B. Olesen, J.S. Clayton, C. Lambertini, H. Brix, B.K. Sorrell. 2012. Growth and morphology in relation to temperature and light availability during the establishment of three invasive aquatic plant species. Aquatic Botany 102: 56-64.
Roth, N.C and A.M. Pregnall. 1988. Nitrate reductase activity in Zostera marina. Marine Biology 99: 457-463
Sagrario, M.A.G.; E. Jeppesen, J. Goma, M. Sondergaard, J.P. Jensen, T. Lauridsen, F. Landkildehus. 2005. Does high nitrogen loading prevent clear-water conditions in shallow lakes at moderately high phosphorus concentrations? Freshwater Biology 50: 27-41.
Salisbury,F.B., C.W. Ross. 1992. Plant Physiology 4th Edition. Wadsworth Publishing Company, Belmont CA. 682 p.p.
Schindler, D.W. 1978. Factors regulating phytoplankton production and standing crop in the world’s freshwaters. Limnology and Oceanography 23: 478-486.
Schindler, D.W., and E.J. Fee. 1974. Experimental lakes area: whole lake experiments in eutrophication. Journal of Fisheries Research Board Canada 32:937-953.
Schroder, R. 1979. Decline of reedswamps in Lake Constance. Symp. Biol. Hung. 19: 43-48.
Schuurkes, J.A., C.J. Kok, C.D. Hartog. 1986. Ammonium and nitrate uptake by aqutic plants from poorly buffered and acidified waters. Aquatic Botany 24: 131-146.
Scott, T., G. Means, R. Meagan, R. Means, S. Upchurch, R. Copeland, R. Jones, A. Willet. 2004. Springs of Florida. Bulletin No. 66 Florida Geological Survey, Tallahassee, FL.
Schjoerring, J. K., S. Husted, G. Mack, M. Mattsson. 2002. The regulation of ammonium translocation in plants. Journal of Experimental Botany 53 (370): 883–890.
Sickman, J.O. A.R. Albertin, M.W. Anderson, A Pinowska and R.J. Stevenson. 2009.A comparison of internal and external supply of nutrients to algal mats in two first magnitude springs in Florida. Journal of Aquatic Plant Management 47: 135-144.
Smolders, A.J., C.D. Hartog, C.B. Gestel, J.G. Roelof. 1996. The effects of ammonium on growth, accumulation of free amino acid and nutritional status of young phosphorus deficient Stratiotes aloides plants. Aquatic Botany 53: 85-96.
Smolders, A.J., M.C. van Reil, J.G.M Roelofs. 2000. Accumulation of free amino acids as an early indication for physiological stress (nitrogen overload) due to elevated ammonium levels in vital Stratiotes aloides L. stands. Archeif fur Hydrobiologie 150: 169-175.
Smolders, A.J.P, R.J.J. Hendriks, H.M. Campschueur, J.G.M Roelofs. 1997. Nitrate induces iron deficiency chlorosis in Juncus acutiflorus. Plant and Soil 196: 37-45.
Stevenson, R.J., A. Pinowska, Y. Wang. 2004. Ecological condition of algae and nutrients in Florida Springs. Florida Department of Environmental Protection, Tallahassee, FL.
Stevenson, R.J., A. Pinowska, A. R. Albertin, J.O. Sickman. 2007. Ecological condition of algae and nutrients in Florida springs: the synthesis report. Florida Department of Environmental Protection, Tallahassee, FL.
Sturgis, R.B., L. Murray. 1997. Scaling of nutrient inputs to submersed plant communities: temporal and spatial variations. Marine Ecological Progress Series 152: 89-102.
Talling, J.F. and J. Lemoalle. 1998. Ecological Dynamics of Tropical Waters. Cambridge University Press. Cambridge UK 455pp.
Turpin, D.H. 1991. Effects of inorganic N availability on algal photosynthesis and carbon metabolism. Journal of Phycology 27: 14-20.
Ulrich, K.E. and T.M. Burton. 1985. The effect of nitrate, phosphate, and potassium fertilization on growth and nutrient uptake patterns of Phragmites australis (Cav.) Trin. Ex Steudel. Aquatic Botany 21:53-62.
Wagner, G. C., 1979, Content and vacuole/extravacuole distribution of neutral sugars, free amino acids, and anthocyanin in protoplasts, Plant Physiol., 64:88.
Wang, Y., G. Gao, B. Qin, X. Wang. 2012. Ecophysiological and anatomical responses of Vallisneria natans to nitrogen and phosphorus enrichment. Knowledge and Management of Aquatic Ecosystems 405, 05.
Wetzel, R.G. 2001. Limnology: Lake and River Ecosystems 3rd Ed. Academic Press, San Diego. 1006 pp.
Weyhenmeyer, G.A., E. Jeppesen, R. Adrian, L. Arvola, T. Blenckner, T. Jankowski, E. Jennings, D. Straile. 2007. Nitrate depleted conditions on the increase in shallow northern European lakes. Limnology Oceanography 52: 13346-1353.
Zimmerman, R.C., R.D. Smith, R.S. Alberte. 1989. Thermal acclimation and whole-plant carbon balance in Zostera marina L. (eelgrass). Journal of Experimental Marine Biology and Ecology 130: 93-109.